免疫组化染色结果支持低分化肝细胞癌伴坏死。Bcl-2(局灶+),CD31(部分+),CD34(—),Ki-67(+ 25-50%),S-100(-),Vimentin(-),CD117(-),CK(+),SM-MHC(-),AFP(-),Hepa-l(-),Melan-A(-),HMB45(—),GPC-3(部分+),Arg-1(+),TFE-3(-)。
在细胞凋亡的相关调控基因的研究方面,Bcl-2基因是目前研究的最深入、最广泛的凋亡调控基因之一。Bcl-2基因最初是在非霍奇金滤泡状B细胞淋巴瘤中分离出来的,它是在14号与18号染色体易位的断点上被发现的。Bcl-2基因编码一个25-26KD的蛋白,其C端的21个疏水氨基酸组成一个延伸的链状结构。这个链可以插到细胞的膜结构中,这一结构特点与Bcl-2调节细胞凋亡的方式和能力非常有关。已经证实Bcl-2存在于线粒体外膜、核膜和内质网膜上。根据对Bcl-2家族成员蛋白的结构研究,发现其共同点是在蛋白链上都存在三个结构基序,分别称为BH1、BH2和BH3结构域。和Ced-9一样,Bcl-2基因属于一类新的癌基因家族成员,通过有效抑制许多不同类型细胞中许多不同类型的凋亡刺激诱导的细胞凋亡,延长细胞活力而发挥其生物学作用,对细胞周期的进程不发生影响。这说明它在细胞凋亡调控机制中起着十分重要的作用,可能是许多因子作用的共同分子基础。但是Bcl-2基因如何抑制细胞凋亡的机制尚不清楚。最近发现Bcl-2可与细胞中的bax蛋白质结合构成不同的二聚体(Bcl-2/ Bcl-2、Bcl-2/bax、bax/bax),通过它们之间的不同比例来调节细胞凋亡。用EB病毒来感染细胞后可使细胞发生永生或生存期延长,这是因为EB病毒促使Bcl-2表达的结果。应用原位杂交和免疫组化方法对鼻咽癌组织和体外培养的细胞进行研究发现,鼻咽癌组织及其体外培养细胞出现bcl-2的过表达。
Bcl-2基因家族是目前广泛研究的一类细胞凋亡相关基因,其表达和调控是影响细胞凋亡的关键因素之一,在细胞凋亡信号转导途径中发挥质量要作用。通过上调促凋亡的Bcl-2的基因表达,从而在转录水平调节凋亡相关蛋白的表达以影响凋亡的发生。Bcl-2家族成员都含有1-4个Bcl-2同源结构域(BH1-4),并且通常有一个羧端跨膜结构域。其中BH4是抗凋亡蛋白所特有的结构域,BH3是与促进凋亡有关的结构域。根据功能和结构可将Bcl-2基因家族分为两类,一类是抗凋亡的(anti-apoptotic),如:Bcl-2、Bcl-xl、Bcl-w、Mcl-1;一类是促进凋亡的(Pro-Apoptotic),如:Bax、Bak、Bad、Bid、Bim,在促凋亡蛋白中还有一类仅含BH3结构,如Bid、Bad。目前已经在哺乳动物、线虫和细菌中发现并鉴定出了bcl-2家族的20余种蛋白质,根据它们在细胞凋亡中的作用可分为两类:一类是抗凋亡蛋白,包括Bcl-2、Bcl-xL、Bcl-W、Mcl-l、AI、Boo和Ced-9等十余个成员;另一类是促凋亡蛋白,包括Bax、Bak、Bcl-xS、Bid、Bad和Egl-l等十余个成员。Bcl-2是迄今研究得最深入、最广泛的凋亡调控基因之一,编码产物Bcl-2蛋白可抑制细胞凋亡,参与细胞增殖与凋亡动态平衡的调控。
CD31又称为血小板-内皮细胞粘附分子(Platelet endothelial cell adhesion molecule-1 ,PECAM-1/CD31),分子量为130kDa,其结构属于免疫球蛋白超家族成员[1],在清除体内老化的中性粒细胞过程中发挥重要作用。CD31存在于血小板、中性粒细胞、单核细胞和某些类型的T细胞表面,以及内皮细胞间紧密连接处。很可能参与白细胞的迁移、血管生成和整合素的激活。
组织分布
CD-31 通常位于血管内皮细胞、 血小板、 巨噬细胞和kuffer细胞、 粒细胞、 T / NK 细胞、 淋巴细胞、 巨核细胞、 破骨细胞、 嗜中性粒细胞。在某些肿瘤也发现CD31的表达,如上皮样血管内皮瘤、 上皮样肉瘤样血管内皮瘤、 其他血管肿瘤、 组织细胞性恶性肿瘤和浆细胞。而在某些肉瘤很少发现,如卡波氏肉瘤[2]。
免疫组化
在免疫组化中,CD31 是主要用于证明内皮细胞组织的存在,用于评估肿瘤血管生成,这可能意味着一个快速增长的肿瘤的程度。恶性血管内皮细胞通常也保留抗原,所以,CD31 免疫组织化学方法还可以用于证明血管瘤和 血管肉瘤。它还可证明在小淋巴细胞和淋巴细胞性淋巴瘤,但CD31并非他们特异的标志分子[2]。
是一种新的、激活后表达水平不发生明显变化的抑制细胞诱导亚群的表面标记。从外周血新鲜分离的CD4细胞中,CD31McAb主要与CD45RA亚群反应,对B细胞合成IgG辅助作用不明显,对ConA和自身MHC(自身MLR)反应较为敏感;而CD31-的CD4细胞群中,发现有大量辅助B细胞合成IgG的活性和对某些抗原刺激的回忆反应。CD45RA 的CD4细胞大激活后,尽管细胞表面丢失CD45RA,但表面CD31的表达仍不发生明显变化;而CD45RO CD45RA-的CD4细胞激活后不能获得CD31表达。由于CD31在CD4细胞激活后仍不变化,对于鉴别抑制细胞诱导亚群和辅助细胞诱导亚群是一种有用的标志。
许多粘附分子如CD11a/CD18(LFA-1)LFA-3,CD2和CD29(VLAβ链)主要表达在CD45RO T细胞表面。而CD31则表达在CD45RA CD4细胞表面。抗CD31McAb作用于naiveT细胞能触发其VLA-4介导的粘附作用。内皮细胞表面CD31及其配体与T细胞表面CD31及其配体相互作用很可能触发整合素介导的粘附作用。CD31如何参与CD45RA CD4 T细胞功能以及诱导抑制性T细胞产生还有待进一步研究。
增殖细胞核抗原Ki-67是一种与细胞周期相关的蛋白质。
Ki-67抗原是目前多种恶性肿瘤尤其是乳腺癌研究中的热门生物指标,可以通过Ki-67抗原的检测,了解恶性肿瘤的细胞增殖活性。
肿瘤增殖Ki67表达范围覆盖除G0 期以外的各增殖周期细胞。研究结果提示PCNA 和Ki67 均可较好地用于检测乳腺癌组织中的增殖细胞,二者基本可以互为代替使用。由于Ki67 在G0 期以外各增殖周期均有表达,更具可用性。两者具有一定的互补性。Ki-67在低分化腺癌组织中的表达较在中高分化腺癌组织的表达明显升高,表明Ki67 染色阳性程度与组织学分级的相关性。表明Ki67表达与乳腺癌发生、发展有关,是一个不良预后因素,对乳腺癌的诊断治疗及预后评价有重要的参考价值。 ki67是增殖指数,越高,肿瘤生长越快,恶性程度越高,一般良性肿瘤ki67小于3%。
CK角蛋白,一般在上皮组织内存在也是上皮性肿瘤,就是癌的标记物,一般癌都是阳性的。
GPC-3在HCC患者血清中中特异高表达,可作为一肝癌血清标志物;其与AFP联合检测可有效的提高肝癌的诊断率;血清中GPC-3与肝癌的临床分期、肝外转移、术后复发及血清AFP水平等密切相关,可作为监测肝癌愈后及预防复发的指标。GPC-3与IGF-Ⅱ基因在肝癌组织异常高表达,可促使肝癌细胞增殖,两者可能在HCC的发生、发展和转移中起重要作用;它们作为分子靶标在肝癌的基因诊断和治疗中的作用尚待进一步探讨。
|
 4年3个月,肺腺癌,五线治疗,依沃西
大家好,特此求助:我爸已坚持抗癌4年3个月,现在替雷丽珠单抗已无效,在经历完33次放
4年3个月,肺腺癌,五线治疗,依沃西
大家好,特此求助:我爸已坚持抗癌4年3个月,现在替雷丽珠单抗已无效,在经历完33次放
 从胃炎到胃癌,身体的求救信号你读懂
作者:Tony
每年的11月是我国的“胃癌防治宣传月”。根据 2022 年全球癌症报告显示,
从胃炎到胃癌,身体的求救信号你读懂
作者:Tony
每年的11月是我国的“胃癌防治宣传月”。根据 2022 年全球癌症报告显示,
 李峻岭教授、王军教授:MET异常肺癌
整理者:雨过天晴审核人:鹰版为帮助MET基因异常等少见靶点NSCLC患者解决在治疗过程中
李峻岭教授、王军教授:MET异常肺癌
整理者:雨过天晴审核人:鹰版为帮助MET基因异常等少见靶点NSCLC患者解决在治疗过程中
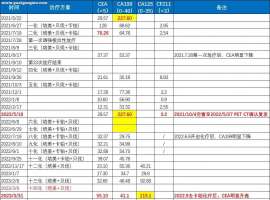 同步放化疗结束3年,化免联合后单免
恳请各位帮忙指导!
2021年7月开始根治性调强放疗,总计33次。近4年治疗方案较简单,
同步放化疗结束3年,化免联合后单免
恳请各位帮忙指导!
2021年7月开始根治性调强放疗,总计33次。近4年治疗方案较简单,
 4个方法助力肺癌患者实现“呼吸自由
作者:王川
清晨,醒来了,却好像没醒透——胸口闷得像压了座山,每一次呼吸都是挣扎
4个方法助力肺癌患者实现“呼吸自由
作者:王川
清晨,醒来了,却好像没醒透——胸口闷得像压了座山,每一次呼吸都是挣扎



 显身卡
显身卡


