| |
| |
| |
| |
 母亲晚期肺癌跨越11年了!我们的5点
讲述者:小白兔整理者:pear
2014年母亲确诊晚期肺腺癌,一路摸索抗癌前行,幸得病友
母亲晚期肺癌跨越11年了!我们的5点
讲述者:小白兔整理者:pear
2014年母亲确诊晚期肺腺癌,一路摸索抗癌前行,幸得病友
 根治性放疗结束后,免疫维持两年能不
家父2022年12月份确诊右肺上叶肺鳞癌3A期,纵膈淋巴结转移,一直在复肿治疗,先新辅助
根治性放疗结束后,免疫维持两年能不
家父2022年12月份确诊右肺上叶肺鳞癌3A期,纵膈淋巴结转移,一直在复肿治疗,先新辅助
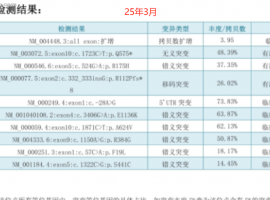 无靶点 her2扩增 二线白紫耐药后怎么
家父今年62岁,22年11月份颈部淋巴结穿刺,确诊肺低分化腺癌,分期是TxN3M1,基因检测
无靶点 her2扩增 二线白紫耐药后怎么
家父今年62岁,22年11月份颈部淋巴结穿刺,确诊肺低分化腺癌,分期是TxN3M1,基因检测
 卡度联合安罗,谁让肿瘤缩小了
肺低分化腺,伴肉瘤样。卡度尼利单抗联合安罗替尼。两个疗程复查显示肿瘤缩小,胸膜增
卡度联合安罗,谁让肿瘤缩小了
肺低分化腺,伴肉瘤样。卡度尼利单抗联合安罗替尼。两个疗程复查显示肿瘤缩小,胸膜增
 褚倩教授、范云教授:肺癌MET变异检
整理者:雨过天晴审核人:鹰版为帮助MET基因异常等少见靶点NSCLC患者解决在治疗过程中
褚倩教授、范云教授:肺癌MET变异检
整理者:雨过天晴审核人:鹰版为帮助MET基因异常等少见靶点NSCLC患者解决在治疗过程中




 显身卡
显身卡